No paleontologist who has studied the flora or fauna of the Comanchean deposits in central Texas has ever reported a human footprint in these rocks. Yet, for decades, the existence of human footprints in these Cretaceous rocks has been touted by creationists as convincing evidence against the geological time scale and evolution. Creationists claim that evolutionists really know that there are human footprints there, as do the local folk. At the very least, they say, Roland T. Bird knew; according to the creationists he practically admitted it in the pages of Natural History and only later attempted to cover it up (Whitcomb and Morris, 1961; Wilder-Smith, 1968). They agree that some tracks were carved in the 1930s to sell to tourists. But they insist that genuine human footprints also exist there (Bartz, 1982a; Beierle, 1980; Dougherty, 1971,1982; Fields, 1980; Gish, 1977; Morris, 1976, 1980; Taylor, 1968, 1970a, 1970b, 1971). Most embrace the dogma set forth in 1950 by Clifford L. Burdick in "When GIANTS Roamed the Earth" that all paleontologists who had visited Glen Rose and surrounding areas for the past half century had been blinded to the truth by their dogmatic belief in evolution! Supposedly, no evolutionists dared admit that the mantracks were real, for mantracks in Cretaceous rock would turn the geological timetable topsy turvy and make a mockery out of evolution. Wilder-Smith (1968:137) put it this way: "It is quite interesting to see what is done in scientific circles with such awkward observations as contemporaneous dinosaur and mantracks. First of all both kinds of tracks were duly reported, but it was suggested by Dr. Bird, who first found them, that either the man tracks or the dinosaur tracks must have been falsified, because according to theory, the two could not exist together!" According to Wilder-Smith (1968: 297), "The giant human tracks have perfectly clear toe, heel and arch imprints. . . . pure theoretical prejudice prevented Dr. Bird from recognizing some exceedingly important geological evidence [against evolution]."
How Can We Tell a Human Footprint?
One of those curious twists of fate is that those very mantracks that led Burdick to write "When GIANTS Roamed the Earth" and that led Whitcomb and Morris (1961) and then Wilder-Smith (1968) to ridicule Roland T. Bird are now recognized as fakes by most creationists (e.g., Morris, 1980). Yet photographs of them still appear in creationist literature. To the human anatomist or physical anthropologist, their fakery is immediately apparent. Most people, however, do not have the knowledge of human anatomy necessary to diagnose the mistakes.
Human footprints differ depending on whether: 1) they are made on hard or soft surfaces; 2) the individual who made them had a normal arch or was flat-footed; and, of course 3) the individual was moving, and at what speed and gait. These conditions might seem to render it difficult to recognize a genuine human footprint, but instead they often facilitate identification, because many features must be right; and conversely many features can be wrong and thus betray spurious human tracks. If, for example, the distance between tracks of a certain length is wrong for a human strider, one can reject a human attribution. What makes track identification difficult is that when tracks are made in soft, wet sediment, mud flowing back into them may obscure anatomical features.
On a nonyielding surface, a human footprint assumes an hourglass shape because the foot bones that articulate with the heel and ankle bones are bound together by strong plantar ligaments to form an arch. One can actually trace an arch in two directions: across the foot and along the long axis of the foot. The arch is high in the middle of the foot, and higher on the inside than on the outside edge. Strong, tight ligaments cause the human foot to behave like a resilient strut; ligaments absorb the compressive stresses that are transmitted to the foot in walking, so the foot muscles have less work to do in resisting these stresses.
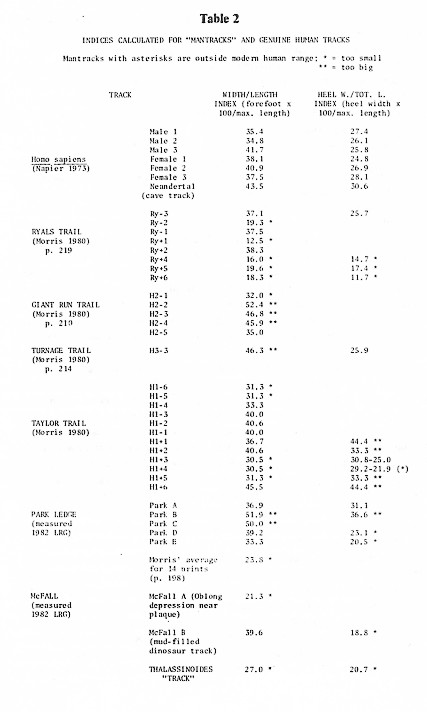
This is why, when a person wets the soles of his or her feet and walks across a hard floor, not all of the sole "prints." Just how much will contact the floor depends on the arch's strength. (Compare the contact surfaces made by individuals with varying degrees of normal to flat-footedness, Figure 1.) Note that contact is always made by the heel (which strikes the substrate first in normal walking), the outside of the foot, the ball of the foot (the pad under the far or "distal" ends of the bones called "metatarsals"), and the big toe or "hallux." These contact surfaces reflect the normal walking cycle (see Figure 2): "Heel strike" first, followed by "midstance" (when the weight is transferred along the outside to the ball of the foot as the pelvic muscles contract to shift the weight of the body over the supporting leg), and finally "push-off" or "toe-off" (when a person propels himself or herself forward by pushing against the substrate with the big toe). Even on a hard surface, one can see the imprint made by the heel, the outside of the foot, the ball, and the big toe. The little toes typically leave only slight marks on a hard floor, not merely because the pads of the little toes are smaller than the pad of the big toe but because they do not bear the weight of the body during push-off.
Notice also that the lateral toes of the foot possess three small bones (called phalanges) whereas the hallux or great toe possesses only two, bigger, phalanges. During push-off the little toes curl up (notice how the joints between the three phalanges make this possible) while the big toe flattens. Only the pads of the little toes contact the floor, and the marks they leave are separated from the mark made by the ball of the foot by a narrow space for the upwardly flexed first two phalanges.
If a person walks across a medium such as wet sand, soft enough to yield to pressure but not so wet that the depressions won't hold after his or her feet are withdrawn, the tracks will exhibit another characteristic shape. They will lack an hourglass outline but their depth will vary from great at the heel and ball to shallow in the arch area. The ball will have made a distinct depression deeper on the inside than the outsidea record of the way pressure shifted just prior to push-off. Mud will have oozed into the curl of the little toes, forming a slightly raised line separating the imprints of the pads of the little toes from the ball. In contrast, the depression made by the great toe will show greater continuity with the ball. In short, the rolling stride of humans produces distortions on a yielding surface that show exceedingly well the anatomical features of the human foot and the characteristics of human striding.
The consistency of the supporting surface also affects stride length. A person's stride is impeded on mud because the degree to which foot pressure imparts motion to the body during push-off is proportional to the counterpressure of the supporting substrate. If the foot slips or sinks in soft mud, more pressure must be applied in order to achieve even slow progress, and the gait becomes relatively inefficient, the stride relatively short.
Anyone can create footprints that do not look terribly human by walking in an awkward manner, e.g., without "pushing-off" with the great toe. This manner of bipedal walking is actually characteristic of some animals such as bears and great apes which occasionally move bipedally (on two legs) but do not have a "rolling" stride. Unlike bears and apes, humans roll their weight from the heel to the big toe, producing an unusually long and efficient stride. If one fails to roll in this manner, stride length will shorten considerably. But we needn't expect that humans walked in such an awkward fashion very often. Even the ancient human footprints preserved in volcanic ash at Laetoli, Tanzania, show heel, arch and ball impressions (Leakey, 1979; Leakey and Hay, 1979; White, 1980), and although the structure of the early australopithecine foot (known from Hadar, Ethiopia as "Lucy") exhibits some clearly ape-like as well as modern human-like features, it also demonstrates that some human bipedal adaptations had evolved by well over three million years ago (Johanson and Edey, 1981). (See Susman, 1983; Stern and Susman, 1983; Susman and Stem, n. d.; Suwa, n.d.; Latimer, n.d.; Gomberg and Latimer, n.d.; and Gomberg, 1984 for discussions of the complex intermediate morphological adaptations of Lucy's foot and their implications for behavior.)
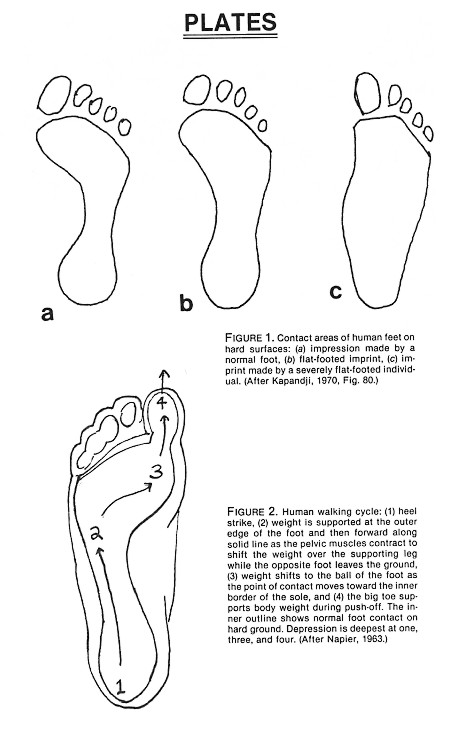
We will examine relative stride length when we look at alleged series of human footprints in Cretaceous rocks. But first we should dispose of the more obvious mantrack forgeries such as the pair of mantracks that so impressed Clifford Burdick thirty-five years ago (Burdick, 1950; see also Figure 11 in Whitcomb and Morris, 1961; Figure 9 in Wilder-Smith, 1968; and Figure 120 in Wysong, 1976). These were the very mantracks that Bird discovered in an Indian curio shop in Gallup, New Mexico, and that initially led him to the Glen Rose area in 1938 (Bird, 1939; Godfrey, 1981).
Artificially Chiseled Giant Mantracks
Burdick's prized mantracks exhibit a suite of anatomical errors (Figure 3). They are poor representations of the modern human footprint, even poorer representations of what giant human footprints might look like, and especially bad representations of what such footprints made in soft mud might look like.
First, the toes are too longfar too long, the big toe far too narrow. The "ball" of the foot is too wide, too far forward, and too deep. It shows no evidence of roll toward the inner edge. The little toes seem to have been carved by someone looking at the top rather than the sole of a human foot. They are chiseled on an unnaturally raised plane; they are also artificially fanned. The heel is too narrow, the forefoot too wide, so that while the relative proportion of forefoot width to total length falls marginally within the range of human variation, the relative proportion of heel to forefoot width is far, too low. The result is an exaggerated hourglass shape. The artisan was apparently trying to reproduce the imprint a modern human foot (with a normal raised arch) makes on a hard surface. The carver did not succeed at that, and certainly failed to produce a facsimile of a human footprint in soft mud where the hourglass outline disappears, and the arch, however strong, becomes a shallow depression. Furthermore, a giant bipedal animal would be very unlikely to have an arch; it would be too heavy.
Now, contrary to creationist claims, there is no fossil evidence of an ancient "race" of giants. Recently Richard Leakey and Alan Walker discovered a "tall" 1.6 million-year-old Homo erectus; that individual, nevertheless, falls within the modern size range. The much later Neanderthals were robust, but they were not terribly tall. Modern humans range greatly in size, from the Ituri Pygmies of Zaire to the Nilotic people of Sudan. Abnormal giantism sometimes occurs, often associated with painful swelling as well as shortened lifespan and abnormally shortened stride. Nevertheless, an anatomist can tell you what the foot of a "normal" giant human might look like, since biomechanical rules of scaling allow us to make predictions even for hypothetical, non-existent, beings. These predictions hold for real footprints made by the more robust Neanderthals and they allow us to understand the shape of the feet of large mammals that occasionally walk bipedally, such as gorillas or bears. In any case it is abundantly clear that a 12 to 16 foot tall Adam (Dougherty, 1978; Burdick, 1950; Baugh, 1983b) would make a footprint nothing like a scaled-up version of a modern human's.
Let's briefly consider the constraints of size on a bipedal animal's foot anatomy and on its locomotor pattern. There is an upper limit to the size of a striding bipeda limit beyond which the characteristic "roll" from heelstrike to toe-off becomes impossible. This is because the human sort of locomotion depends on the big toe supporting the weight of the body during toeoff. Individuals must be light enough, and the hallux strong enough, so that the big toe will not break from the compressive load and bending stress to which it is subjected in walking. If an exceedingly large human with perfectly "normal" walking gait existed, his or her feet would have had to exhibit a set of allometric structural alterations. Specifically, such a person would have possessed relatively wide feet with weakly developed arches, relatively short little toes and relatively wide and short great toes. The latter characteristic is especially important if a rolling stride is to be preserved.
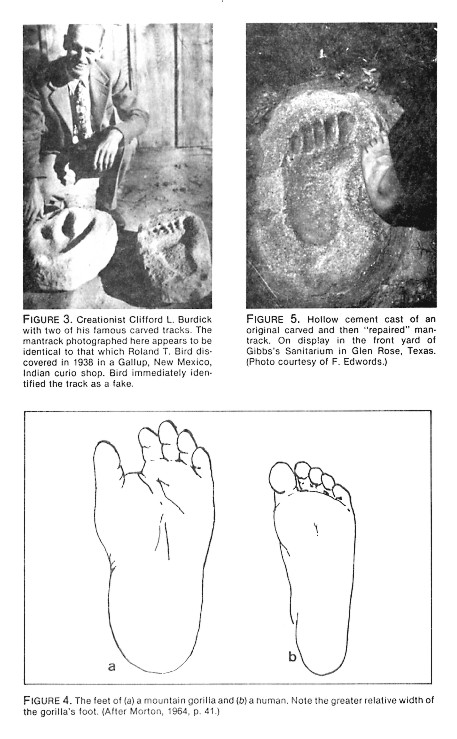
The size limit for a humanlike rolling stride may in fact be exceeded by animals as small as gorillas or bears. Figure 4 compares the outline of a human foot with that of a gorilla. Gorillas weigh approximately twice what humans weigh; male gorillas may exceed 500 pounds. Gorillas normally walk on all fours, but occasionally they move bipedally. When they do, they do not use the big toe for push-off. These so-called non-striding bipeds have relatively shorter cycles than do humans (that is, their step is short relative to hindlimb length or body weight) because they must lift up their feet and place them down flat, without rolling off the big toe. The biomechanical properties of a striding foot are fundamentally distinct from those of a non-strider, and these differences are reflected in differences in the morphology of the foot and in the imprint of the foot on both yielding and non-yielding surfaces. A gorilla's foot makes a relatively wide and short impression. The toes curl to the side and make very light imprints. Bear footprints have many similar properties: they are short and wide (with width/length indices in the low 50s rather than the mid-to-high 30s and low 40s characteristic of humans). Because bears do not push-off with their toes, the distances between successive footprints are relatively short.
A humanlike biped capable of making tracks roughly equal in depth to those of a large dinosaur would not be capable of striding in a human fashion; it would weigh too much. The foot would have to be proportionately much wider than the human foot; the imprint would be deepest in back instead of under the ball of the foot, and toe impressions would be relatively light.
The practical jokers who carved the giant mantracks not only did a poor job of reproducing the shape of a human footprint, but they erred in the wrong direction. They failed miserably to compensate for the grotesquely large size of their subjects. The prints deviate from the modern norm in the wrong direction given either: 1) the supposition that they were produced by an animal still capable of modified striding (in a human sense); or 2) the supposition that they were produced by an animal that had exceeded the normal size limits of a humanlike strider, and was using instead a relatively shorter cycle and more flat-footed gait, with greater weight transmission through the posterior part of the foot.
Similar mistakes are exhibited on other chiseled prints, as, for example, another right footprint in Burdick's collection which was figured by Whitcomb and Morris (1961: Figure 10) and used to make the cast which appears in Wilder-Smith (1968: Figure 10). Once again the hallux is far too long given its own width (its width should be increased by about 3/4), and it is about a third too long given the total length of the footprint.
W. Gibbs, the owner of a cement mantrack on the lawn of his Glen Rose sanitarium (Figure 5) says that his mantrack was made by one of the Adams brothers to cover a genuine giant mantrack that had been vandalized by some pranksters and thus needed to be repaired. He says that Adams accurately reproduced the true anatomical features of the original mantrack. Morris (1980) also treats this track as genuine. But its great toe is even more grotesquely elongated than that of Burdick's track, both in relation to its own width and in relation to the maximum length of the footprint.
Footprints in Situ: The Creationists' Own Published Accounts
Since the late 1960s, creationists have attempted to downplay obviously carved specimens, and to concentrate on exposing "fresh" mantracks, in situ. They have published maps and poor photographs of these features (Beierle, 1980; Fields, 1980; Morris, 1980), that should ideally allow one to locate and examine them firsthand. , This is often more difficult than it might seem: First because some of them are erosional features which differ little from the surrounding surfaceonly when they are painted with water can one see their "human" characteristics. Secondly, many of the mantracks reported in the creationist literature have been destroyed by further excavation, direct removal, or erosion. Some are under water most of the time. No casts have been made available to professional human anatomists or ichnologists. However, the creationists' own published field notes, measurements and photographs, however poor, allow us to evaluate their claims.
The creationists' mantrack data are given in Tables 1 and 2. (These measurements were recorded originally in inches or centimeters but are uniformly converted to millimeters here for easy comparison. Stride and pace measurements are expressed here in meters.)
Generally three to five measurements are reported in the creationist literature:
- Width across toes (sometimes "width at center" or just "width"; in the latter case it is unclear where the measurement was taken).
- Length. Presumably maximum length.
- Width across heel. These measurements were inconsistently recordedmost often only by Wilbur Fields.
- Pace or stride. Creationists use the terms "pace" and "stride" interchangeably to mean the distance between one footprint and the next. Thus Wilbur Fields (1980) records "pace" and Morris (1980) records the same measurements as "stride." Morris is technically incorrect. Pace is properly defined as the distance between some fixed anatomical point on one footprint and the same anatomical point on the next footprint (opposite side), whereas stride is the distance between that anatomical point and the same feature on the next track made by the same foot. Thus stride is roughly (but not exactly) twice the length of a pace in an efficient bipedal animal. Note that pace is measured oblique to stride, and in the case of an inefficient biped whose feet are placed wide apart, pace may be far greater than half the stride. For the sake of accuracy, all of the creationist measurements of distance between two successive tracks are here called pace; stride is used as defined above.
- Depth. Depth was not consistently recorded. It should be noted that the depth of an impression depends more on the nature of the substrate than on the weight of the animal. Even a very light animal will make deep impressions in mud of almost quicksand consistency. It is thus useful to know depth only if the approximate weight of the animal making the track is known: then the depth can help the footprint specialist decide whether the track was made on a hard intertidal surface, in soft sediment under shallow water, or in deeper water. Similarly, if the track of an animal of unknown weight is preserved adjacent to the track of an animal of known weight, the relative depths can be used to evaluate the weight of the unknown trackmaker. Thus an alleged mantrack inches from a sauropod track of the same depth would be suspect! Also, mud on a lagoonal tidal flat can vary from wet to firm in short distances, so tracks in a single trackway may vary considerably in depth. This phenomenon was clearly recorded in some tridactyl (three-toed) dinosaur tracks preserved at the McFall site near Glen Rose. Trackways contained both very well and very poorly demarcated dinosaur footprints; poor tracks were formed when the dinosaur's foot sank deep into wet mud. Mud then oozed back into the depressions made by the front three toes when the foot was withdrawn, leaving only fanning up front and a distinct, elongated "heel" to the rear. Actually, the "heel" was produced by the dinosaur's hallux, which points rearward, as it does in modern birds. The resulting elongated depression actually represents only a portion of the dinosaur's foot and can look vaguely humanlike. (Indeed, these marks have been mistaken for mantracks by some creationists, and what is more startling, when these creationists observe good tridactyl dinosaur prints in the same series as their "mantracks," they conclude that the "man" stepped into the dinosaur trackway, or vice versa. It apparently doesn't seem odd to them that their "man" and dinosaur took exactly the same strides and followed exactly the same paths.)
John Morris's (1980) book Tracking Those Incredible Dinosaurs . . . and the People Who Knew Them is usually considered the most authoritative creationist source on Texas mantracks. Morris compiled data, many of which had appeared elsewhere (e.g., Fields, 1980), but he remeasured none of the trackways, even though he claims to have "improved" some of Fields's measurements. Thus one finds that some measurements from different creationist sources agree remarkably well; they are, in fact, the same measurements republished. The differences between them reflect only rounding error that occurs when, for example, Fields's measurements, taken to the nearest centimeter, are expressed to the nearest half-inch by Morris. Striking differences among values for the same features appear when they were measured by different creationists at different times. Morris's "improvements" also result in drastic unexplained changes of some measurements.
Of course even when creationists record such measurements as "width at toes" or "width at heel," it is not because they see actual toe or heel imprints. They are simply measuring the wider and (sometimes) narrower ends of elongated depressions which they take to be toe or heel marks.
How Can We Tell a Human Trackway?
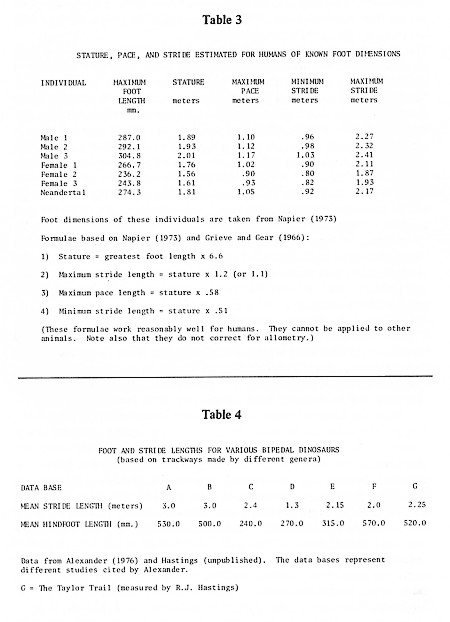
When an animal moves, the impression it makes with its foot depends upon the absolute amount of force transmitted to the substrate and the way that force is transmitted to the substrate. This in turn depends upon the weight of the organism, the structure of the foot, and the gait (walking, running, galloping, etc.). The characteristics of a trackway reflect all of these things, as well as the nature of the substrate. In general, the distance between tracks increases with increasing speed, although the relationships change when the animal shifts gait. Because a number of variables are linked (e.g., stepping frequency, speed, pace length, stride length, foot length, stature), one can make fairly good estimates of unknown values for some of these variables when the values for others are known. Thus, for example, a person with a given foot length will have a comfortable average pace and stride, a minimum pace and stride, and a maximum pace and stride. Because humans are highly efficient bipeds, pace length is always about half the length of a stride, even in slow walking. Minimum and maximum values for pace and stride can be estimated from foot length alone. (See Napier, 1973 and Grieve and Gear, 1966 for discussions of formulae one might use for humans.)
One can use creationist measurements to evaluate the anatomical proportions of alleged mantracks by comparing values obtained for indices of supposed mantracks and actual human footprints. And one can use the measurements to evaluate the plausibility of claims that the trackways were produced by striding humans by comparing measured paces with paces projected for humans having mantrack foot lengths. Some human values are given in Table 3, along with formulas for calculating minimum and maximum stride lengths given observed foot lengths. Let us begin with how we might use such data to evaluate one of the "better" supposed human trackways.
The Taylor Trail
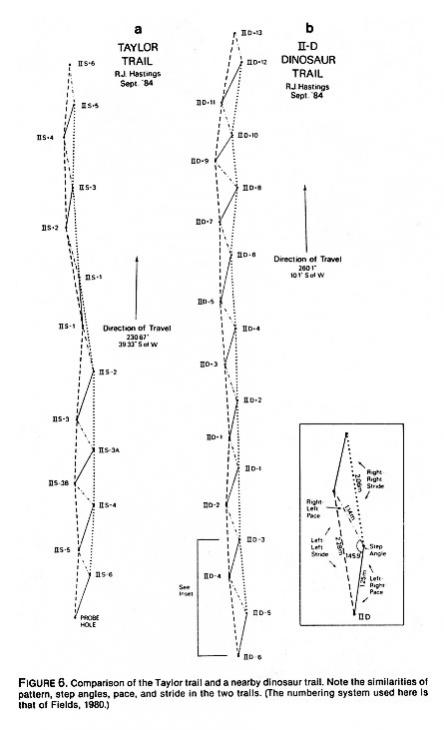
Table 1 reproduces the data in Morris (1980: 206) for the so-called Taylor Trail, a trackway excavated by Stanley Taylor's crew and filmed in Footprints in Stone (Taylor, 1970a). It is still considered one of the creationists' best trails. This is a genuine trackway made by some bipedal animalbut was it human? Taylor measured six tracks in 1970; Wilbur Fields returned to the site in 1977 and produced a second set of measurements on these and several additional tracks in the same series. He retook some of Taylor's measurements (width at "toes" and maximum foot length), and added others that Taylor had not recorded, such as pace length and width across "heel." (Recently a member of our team, Ronnie Hastings, returned to the site when the river was dry and was able to obtain a complete set of measurements for the Taylor trail. Some of these are included in Table 1, along with the available data of both Taylor and Fields. Hastings's measurements will be discussed following an analysis of the creationists' own results.)
It is immediately apparent that there is no agreement between the measurements taken by Fields and by Taylor for the same tracks. Morris acknowledges this fact, and then offers an explanation:
Fields measures the average length at about 16 inches while Taylor found that the best prints, the ones with no evidence of slippage, averaged about 10 inches. Erosion has taken a deadly toll. (Morris, 1980: 207)
This is a rather remarkable explanation, since it seems to offer both erosion and slippage as explanations for the discrepancies between the measurements taken by Taylor and, seven years later, by Fields. Presumably erosion would have taken place between 1970 and 1977; but slippage would have been present (if at all) in both 1970 and 1977, and no anatomist would include slippage marks in measuring foot length. Given the discrepancies, one must conclude that either Taylor or Fields (or both) didn't know what he was measuring. If we accept Morris's remark about erosion we might be tempted to accept his ten inch (254 millimeter) foot length for the individual who produced this trackway. This is far smaller than even the smallest measure recorded by Fields (279.4 mm), and about half Fields's longest measurement (457.2 mm). Fields's average footprint length was 406 mm and his average width at "toes" was 140.8 mm, while Taylor's averages were 314.3 and 99.5 mm. Morris is asking us to believe that erosion lengthened and widened the tracks in seven years by, on the average, more than 50%.
But there are bigger problems. If we decide that the actual length of the trackmaker's foot was Morris's 254 mm (and if we assume that the trackway was produced by a striding human), we obtain estimates of stature of 1.68 meters (5 feet 5 inches), minimum and maximum stride of .86 and 1.88 meters, and maximum pace of .97 meters. This maximum pace is actually far smaller than the paces recorded by Fields (see Table 1). Fields recorded a series of paces ranging between 1.12 and 1.42 meters and averaging 1.31 meters. These walking paces simply could not have been made by a 5 foot5 inch (1.68 m) human. They are far too big. They are even too big for a seven-foot tall human. Moreover, Fields recorded some paces of 2.03 and 2.31 meters (but see below). The latter is the maximum pace for a thirteen-foot tall human (assuming we can make such an extrapolation at all with no correction for allometry, which is a mistake)! So, if we accept Fields's pace measurements, we have to throw out all of the measurements taken by Taylor and crew immediately after the trail was first exposedbefore, as creationists say, erosion took its toll. Morris's argument that the Taylor Trail was made by a human with 10 inch feet can be refuted by inconsistencies in the creationists' own data set.
There is ample evidence that Taylor didn't know what he was measuring in the first place (how can a single human footprint be 304.8 to 406.4 mm long?). Furthermore, there are internal inconsistencies in the data sets collected by both Taylor and Fields. Fields's lengths vary from 279.4 to 457.2 mm; Taylor's from 228.6 to the vague "330.2 to 406.4 mm." Fields's widths vary from 127 to 165.1 mm; Taylor's from 88.9 to 101.6 mm. If we calculate width/length indices using data collected by either investigator, we find marked changes in foot shape along the trail. The discrepancies are even worse when we compare values for indices based on Fields's data with values based on Taylor's data for some of the same tracks! For example, track H1+1 has a width/length index of 36.7 (Fields) and 44.4 (Taylor). Worse yet, half of the creationist values for this index fall outside the range of normal human variation (this is especially true of those based on Taylor's data), and moreover, they are too low (Table 2). If any of Fields's high footlength measurements are to be believed, we should expect this giant to have had unusually wide feet and thus higher than normal values for its width/length indices.
Hastings's detailed map and measurements of the Taylor trail prove beyond any doubt that these tracks could not have been made by a giant human (Figure 6a). They also serve to demonstrate the enormity of measurement and observational errors made by both Taylor and Fields. First of all, despite the poor quality of the tracks in this trail, some of the tracks exhibit faint claw marks and fan out in the front, as is typical of dinosaur footprints. (Hastings informs me that even more distinct claw marks are visible on some of the tracks in the so-called Turnage trailMorris's "most humanlike" trackway.) Secondly, the Taylor tracks form a pattern that is atypical for humans but not for bipedal dinosaurs: the animal that made them shifted from an irregular somewhat bouncy gait with uneven pace and short stride to an increasingly regular, faster gait. Track breadth narrowed as the animal assumed a more efficient gait; the feet were placed closer to the central axis of the trail and the stride lengthened. Human tracks tend to be very regularly spaced, and the trails consistently narrow. The pattern and step angles of tracks in the Taylor trail show marked similarities to those known to be made by dinosaurs (compare Figures 6a and 6b).
Finally, it should be noted that Hastings's recorded pace, stride, and foot lengths for the Taylor trail fit known values for dinosaurs and not humans. The shape of the trackselongate grooves that are deepest in the centers with some splaying up frontcharacterizes many poor dinosaur tracks and not human tracks. The obvious, most parsimonious, explanation is that this trail was made by a bipedal dinosaur.
Let's Play Hop, Skip and Jump
Footprints in Stone shows creationist Mike Turnage skipping from one mantrack to another in a trail. The point of this frolicking seems to have been to raise the question: If a modern man can skip from one mantrack to the next, wouldn't it have been a snap for the human giants of olden days to have made them? How could a modern human so easily retrace the steps of dinosaurs?
In fact, modern humans can hop, skip and jump from one dinosaur track to another in a series. First of all, because they use a rolling stride, humans take relatively long paces for their body size. Secondly, people can s-t-r-e-t-c-h in an awkward manner and almost double their pace for at least one or two steps. Furthermore, while dinosaurs were apparently capable of rapid movement (Halstead, 1982), they often moved slowly (Alexander, 1976; Thulborn, 1982), taking short steps relative to foot length (or, especially, body mass). And these Texas dinosaurs were moving on soft mud; humans retracing their steps today are moving on hard limestone. If there were humans walking on the mudflats alongside these dinosaurs, they would have taken shorter strides than we can take on the now-hardened rock.

Normally, adult humans walking comfortably take paces of about .6 to .9 meters. Comfortable strides will be about twice that. Slowing down or speeding up, the range of possible strides is much greater (see Table 3). A small female may exhibit a normal range (minimum to maximum) in stride from about .8 to about 1.8 meters. (Note the possible overlap between normal pace and short stride lengths.) A tall male may exhibit a normal range in stride length from about 1.0 to 2.25 meters. One exceptionally tall (6 foot 10 inch) male whose pace and stride we measured exhibited a normal, comfortable stride of about 2 meters; his comfortable pace length was consistently 1 meter long. Using his foot measurements, his minimum stride length can be calculated at a little more than 1 meter, his maximum stride length at about 2.3 meters, and his maximum pace at about 1.2 meters. He was capable of stretching his leg to reach a "super-pace" of 1.7 metersi.e., 1.7 meters in one stretch-step! If dinosaur tracks in a series are 1.3 meters apart, one can see how people even shorter than this man might easily reach them by stretching. But they could not keep this up in a normal walk. And they would have extra difficulty doing so in mud.
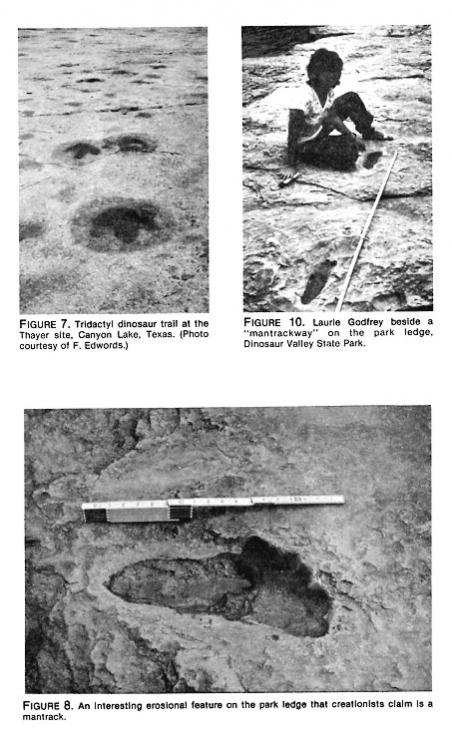
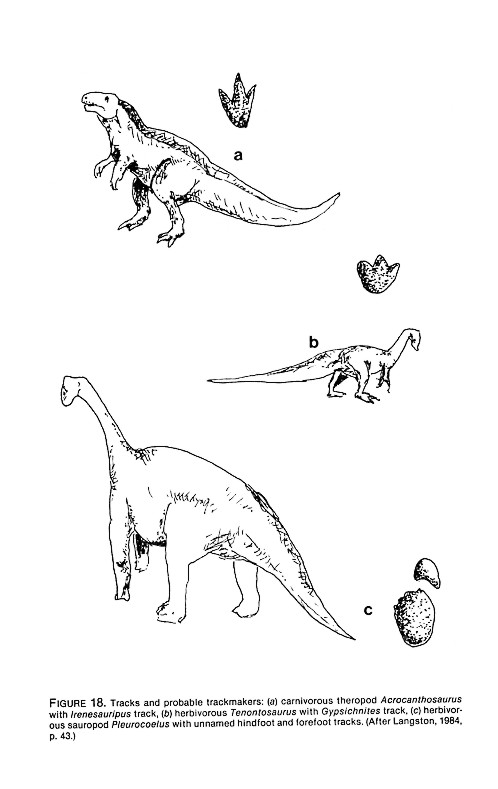
We measured tridactyl dinosaur strides, paces, and foot lengths at the Thayer (New Braunfels) site, and extensive maps of this site plus detailed measurements have been made by paleontologists Wann Langston, Jr., and James Farlow (Figure 7). Foot and stride lengths of other bipedal dinosaurs have been published (see Table 4). New measurements for Texas bipedal dinosaur trackways are currently being compiled by James Farlow.
When the clarity of tracks allows it, precise measurements of pace, stride, lengths, and width can be made. But one must pay careful attention to anatomical landmarks to obtain measurements that can be usefully compared. For example, an ornithopod footprint shows three forwardly facing toes. Different "lengths" can be obtained by measuring along each separate toe. One Gypsichnites track, for example, measured 440 mm along the middle toe; 350 and 300 along the lateral toes. Maximum breadth for this particular specimen, across all three toes, was 370 mm. Not surprisingly this large dinosaur had a very high width/length ratio (370/440 or .84). But as is typical of tridactyl dinosaurs, each toe was relatively narrow, and an impression of a single toe gives the dimensions creationists often record as typifying mantracks. Pace lengths also match typical creationist measurements for man-trackways. For example, this particular ornithopod took 11 steps in 15.81 meters.
Pace and stride length changed gradually throughout the sequence, increasing and then decreasing slightly. We measured the following sequence (strides from tracks 1 to 11, i.e., 1 to 3, 3 to 5, 5 to 7, etc.): 3.11 m, 3.18 m, 3.21 m, 3.17 m, 3.11 m. The longest paces were about 1.6 meters. Many trackways at the Thayer site exhibit shorter strides. One dinosaur took 15 steps in 15.1 meters, covering an equal distance in as many steps as is typical of our 6-foot-10-inch human! But then from track 10 to 18 stride length increased gradually from 2.04 meters to 2.55 meters as the animal increased its speed. Its longest stride probably exceeds the limit for our tall human subject, but only by a little bit. Overlap in single measurements is obviously not the whole story. The stride lengths of humans and dinosaurs overlap, but the whole gestalt is not the same. The ornithopod that took roughly one-meter-long steps over a short distance had much larger feet than our human does. And it was clearly capable of taking much longer strides. To test human origin claims, the range and combination of foot plus stride and pace measurements must be checked.
Bipedal dinosaurs varied tremendously in size; some were quite small. Furthermore, many dinosaurs were "semidigitigrade" or "digitigrade"; not all of their foot contacted the ground even in normal walking. Some species left tracks that were 150 to 200 mm long (shorter than those of modem adult humans) and took 1.5 to 1.8 m walking strides. Some species made tracks over 500 mm long and took normal walking strides of about 3 m. (Running dinosaurs took much longer strides relative to their track lengths.) Walking paces of a meter or more are rare for humans and common for large bipedal dinosaurs. Human trackways are narrow; bipedal dinosaur trackways are broad or narrow. One can always speculate that "giant" humans took giant paces, but would they have had feet as narrow as individual dinosaur toes or "heels"? Would their feet have been relatively narrower than those of modem humans? The laws of scaling seem to indicate that this is impossible.
The Ryals Trail
John Morris (1980: 219) published data collected by Wilbur Fields for the Ryals Trail at the McFall-Taylor site (Table 1). These prints are usually under water but were filmed in Footprints in Stone. Morris gives measurements for nine tracks in this series, most taken directly from Fields. Others, which deviate markedly from Fields's primary data, are called "improvements" (e.g., pace length of 1.04 meters instead of 1.75 meters for Ry+4). Minor differences between most of the measures are due to rounding errors. Morris's data are presented in parentheses beside those of Fields.
The Ryals trail is often featured by creationists because it includes a footprint (Ry+2) which is said to show evidence of a human hallux. Although I was unable to examine it firsthand, photographs of it in Footprints in Stone show that it has none of the features of a human great toe impression, and this is, interestingly, confirmed by the creationists' own measurements. The toe imprint is a small hole that is far too small (relative to the total length of the impression) to be even vaguely humanlike. To have modern human toe proportions, this "great toe" should be three times its actual size. In fact, it appears to be a claw mark obscured by sediment infilling. As might be anticipated, there are no impressions for lateral toes adjacent to that of the "great toe."
The Ryals trail exhibits all of the classic problems that characterize creationist mantrackways:
- Measurements such as maximum footlength vary tremendously (in this case, from 350 to 750 mm) indicating at once that the tracks are too poor to exhibit precise anatomical landmarks, or that the creationists had no idea of how to recognize those landmarks, or both. Widths also varied wildly; according to Fields, Ry+2 was almost five times the width of Ry+1 (50 versus 230 mm)! The foot seems to have changed shape as well as length, so Fields felt compelled to measure the width of Ry+2 at the center whereas all other widths were measured at the "toes."
- When measurements are retaken, values reported by different creationists show no agreement (compare the values of 230 and 127 mm. given by Fields and Morris for "width" of Ry+2; compare the pace lengths of 1.75 and 1.04 in given by Fields and Morris for Ry+4; then note that Ry+2 and Ry+4 are, according to Morris, the best of these supposed mantracks).
- The foot proportions vary as much as individual measurements, so that width/length ratios may double or even triple for tracks in a series (compare 12.5 and 38.3; Table 2)! Moreover, a good many of the values for these indices (in this case nine out of thirteen) fall far outside the range of human variation. This is true of the width/length ratio, the big toe length/total foot length ratio, and the ratio of heel width to total length.
- Values for indices deviate in the wrong direction from allometric expectations for a supposed giant human. Thus the markedly deviant values for the Ryals trail indices shown in Table 2 are all too low.
- Pace lengths match those of dinosaurs and are almost universally outside the range of modern walking humans.
- The lengths of the features measured match the lengths of known dinosaur footprints.
- 7) The tracks exhibit none of the tell-tale signs of human footprints-the hallux continuing the ball, the ooze of mud under the toes, the different relative depths of different parts of the imprint.
Dinosaur Valley State Park
Imagine how hard it would be to measure something if nothing was there. Well, all mantracks on the Dinosaur Valley State Park ledge are exactly thatnothing at all. Perhaps I should qualify that statement by saying that they are not tracks; they are elongated erosional grooves with nebulous boundaries (see Figure 8). The "Brontosaurus" and "bear" tracks (Morris, 1980: 156, 228), are also erosional features. Without the creationists' maps and photos we could never have located them. Actually, none of the park's many dinosaur tracks occur on this particular stratum, but it is a popular mantrack site for visitors unwittingly impressed by erosion.
There is ample evidence that creationists have also had difficulty defining borders and locations of these alleged mantracks. As Milne and Schafersman (1983) point out, one print was illustrated by Morris (1980: 229, top left photo) as a definite left, while both Dougherty (1971 cover photo) and Beierle (1980: 32 and 33) describe it as a definite right footprint. Each showed (but in different places) a "big toe" mark. When one looks at these prints without highlighting anything, one sees typical erosional channels and pits. Erosional pits become toes if they happen to be roughly the size of a modem human toe (see Figure 9). Erosion creates a very irregular surface on limestone, and river erosion creates elongated grooves roughly parallel to the direction of flow of the river. Some depressions, of course, will be roughly the length of a modem human footprint. These are the features creationists have identified here as mantracks. They occur "in series" only because the direction of river flow was uniform. As might be expected, these mantracks have a host of things wrong with them. The distances between them ("paces"?) are extremely irregular (measuring in one case 2.18 and 1.40 meters for successive "footprints"). The so-called hallux impression, when present, may be wider than it is long (unlike genuine human big toe prints). Whereas the total lengths of some of these features do not exceed those of modem humans, their width/length indices fall in the range of bears rather than humans; compare width/length values for erosional features B and C with actual human values (Table 2). One would not expect human footprints of these lengths to be so exceptionally wide. Other, longer, features turn out to be considerably narrower, again contrary to expectations for genuine human footprints (see Figure 10). Unlike the toes of the carved mantracks which are too long, these "toes" (actually erosional pits) tend to be too small and especially too short. None of the features show the tell-tale signs of human footprints. So-called "insteps," "heels," and occasional "toe marks" are formed by karren dissolution, erosional undercutting and the creativity of imagination that allows us to see camels in cloud formations.
The McFall Site
Old Emmett McFall's property is the Reverend Carl Baugh's playground. Baugh (1983b) claims to have found 44 human footprints in situ here. While maps and measurements of Baugh's new mantracks have not been published, members of our team were able to visit the site on a number of occasions shortly after fresh mantracks had been exposed. Baugh and members of his crew pointed out track locations to us. A brass plaque commemorating the discovery of Humanus Bauanthropus is located near an elongated groove that Baugh took for a giant mantrack.
None of his mantracks are recent erosional features. They all fall into one of three categories, however:
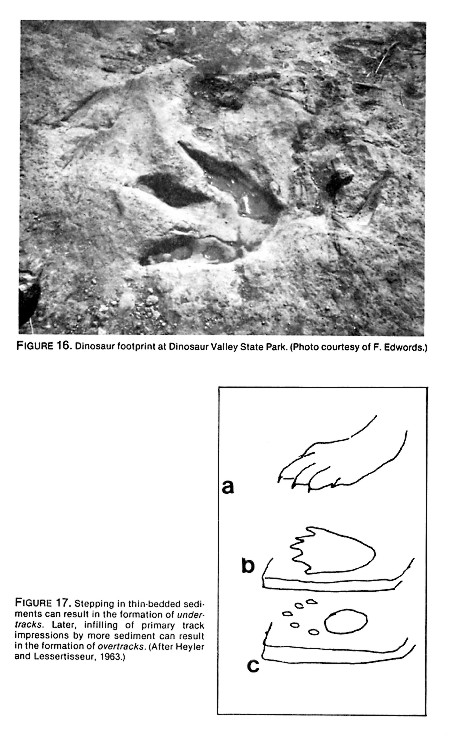
- Clear toe impressions of tridactyl dinosaurs. The limestone beds at the McFall site and other neighboring properties abound with footprints of tridactyl dinosaurs whose tracks have been named Irenesauripus and Gypsichnites (Langston, 1983). Some "three-toed" dinosaurs actually had four toes. The hallux, situated at the rear of the foot, sometimes dragged in mud, creating a distinct heel impression (Figure 11). The depth or even existence of this rear toe impression depends upon how far the dinosaur's foot sank into the mud. If one is not picky about shape, one can imagine that this heel (or rear toe) mark is an entire human footprint. Needless to say, the "toes" of such footprints are obscure as is, indeed, the entire outline. (But Baugh tries to turn this into an advantage by claiming to have discovered proof that the humans and dinosaurs lived together because of the astounding fact that the humans stepped into the dinosaur tracks or vice versa.) Due to the difficulty of finding the front border of these mantracks, lengths of mantracks, three in series, seem to be six inches, ten inches and sixteen inches, for example.
- Poor dinosaur tracks. Other Baugh mantracks are not portions of clear dinosaur footprints, rather they are poor dinosaur tracks that have been obscured by partial infilling of mud. They can be found in series with other much clearer dinosaur tracks. Fanning in the front usually shows the axes of three partially-filled toes (Figure 12). One such mantrack at the McFall site measured 480 mm long, 90 mm at the "heel," and 190 mm at the front where the impression fans out and disappears. It should be immediately apparent that these dimensions match those of many creationist mantracks and fail to match the expected proportions of human footprints (especially giant human footprints). The "heel" is much too narrow; the length too great. There is no ball impression or any other anatomical feature that characterizes human footprints. The so-called "Giant Run" mantracks (Morris, 1980) are all of this character (Hastings, personal communication).
- Some of these depressions are even more obscure. They may be formed when, for example, mudflow completely hides the fanning at the front of a dinosaur footprint leaving only a slitlike impression, or when the dinosaur makes a scoop mark with a foot or a tail that briefly contacts the surface. The oblong depression located near the Humanus Bauanthropus plaque has this character (Figure 11). It scoops up on both sides and is long and narrow. Its length (470 mm) and width (100 mm) match those of nearby clear dinosaur toe prints; these dimensions are certainly wrong for a human footprint, being once again far too long and narrow. The depression is also deepest rather than shallowest in the supposed arch area. The "toes" that some creationists have identified at one end of the impression are actually small and indistinct erosional pits that have none of the characteristics of actual toe marks. One can count five, six, sevenhowever many one wants. One woman, clearly a believer, told us that the big toe must have been in the middle of the "giant human's" foot because the biggest shallow pit happened to be in the middle.
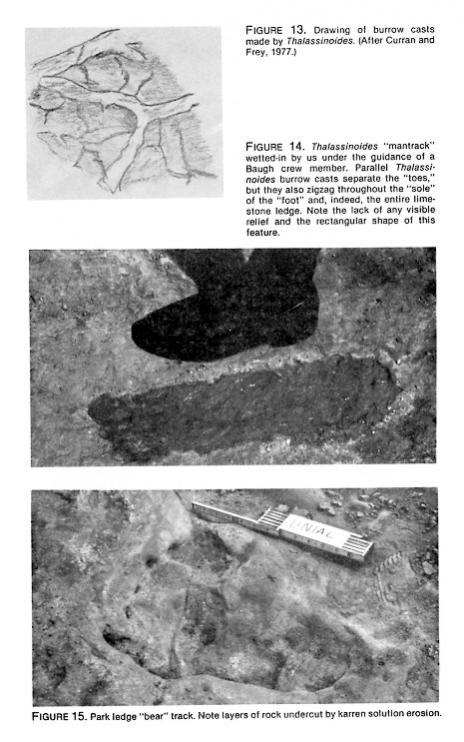
- Thalassinoides. Perhaps the most remarkable mantracks are those Baugh and his crew created out of invertebrate burrow casts of Thalassinoides (Schafersman, 1983). An exposed limestone bed at the McFall site is covered with such burrow casts; they form lattices and ridges that, to the creationists, sometimes separate human toe impressions or saber-toothed cat pads. A member of Baugh's crew showed us the outline of one such mantrack that we were then able to measure (Figures 13 and 14; Table 2). Its "big toe" was 43.6 mm wide and 44.5 mm longroughly square, in other wordsand also far too small for the length of the footprint (450 mm). Its width/length index was too smalloutside the range of human variation. None of the salient anatomical features of genuine human footprints was present; in fact, this footprint had no relief at all, except for the burrow cast ridges that covered its entire surface. Genuine trace fossils? Yes! Genuine mantrack? Definitively, no!
Conclusion
Any claim of human and dinosaur contemporaneity based on the alleged discovery of both kinds of footprints in the same rock deposits will be treated seriously by the scientific community only if it is based upon clear tracks of both. Excellent dinosaur tracks abound in the Cretaceous rocks of central Texas. In contrast, all of the alleged mantracks are miserable. The question addressed in this essay is: how can one recognize a genuine human footprint?
In order to determine that a given depression is a genuine human foot print, we need to understand human footprint anatomy. How do anatomical principles govern variation in foot size and shape? How do the impressions human feet make vary on different surfaces? How is the pace-and-stride pattern humans make constrained by stature and gait? Only after one specifies the predictions of a hypothesis can one determine whether observations fit them. Thus one tests the hypothesis that a given set of depressions is human by specifying and then looking for tell-tale signs of human anatomy and gait.
When we approached the evidence in this way, we found that the alleged Cretaceous mantracks consistently failed the test of human origin but often passed the test of dinosaur origin. Indeed, some were quite clearly portions of dinosaur footprints. Othersthose most responsible for the Paluxy mantrack legendturned out to be inept carvings. Although these were definitely in the minority, had they never existed it is doubtful that creationists would have focused on the Glen Rose area in the first place.
We not only measured many of the trackways that creationists claimed were human, but we also scrutinized the creationists' published data on mantracks. We consistently found these data to be shoddy and, even when taken at face value, to lead to absurd conclusions about the stride length, foot length, and foot shape of the "humans" that presumably made them. The inescapable conclusion is that there is no footprint evidence in Texas supporting the notion of human and dinosaur contemporaneity.