 The topic of the evolution of birds always interests NCSE readers, and so it was not surprising to find so many good questions in our mailbox that we thought it would be interesting to collect them all together and respond in a single column. First, however, I have to apologize to readers and thank John Ruben of Oregon State for pointing out a misstatement on my part. I said that some dinosaurs such as Beipiaosaurus and Sinosauropteryx had beta-keratins in their integumentary coverings (RNCSE 2000; 20 [5]: 28-31). Sorry! Rather, the alvarezsaurid dinosaur Shuvuuia, which is close to oviraptorids, has beta-keratin in the filamentous, featherlike integumentary structures on its head (Schweitzer 2001). More importantly, like birds and unlike other reptiles, these structures lack the alpha-keratins found in reptile scales and all vertebrate skin. Beipiaosaurus and other feathered and "fringed" non-avian dinosaurs are in the same clade of birds and closely related theropod dinosaurs, but, contrary to what I noted, they have not yet been tested for keratins.
They have the same kinds of integumentary structures, which appear to be homologous to the feathers on today's birds as well as to those of ancient birds such as Archaeopteryx and Confuciusornis. Because all these animals have the same kinds of structures, and they are in the same clade of closely related animals, we infer that all these structures lacked alpha-keratins, and that hypothesis must be tested by future discoveries. I apologize for creating confusion: I was thinking of one animal and writing about another, and my fingers got ahead of my brain (or vice versa). For more information on feather biochemistry and evolution see Brush (2001 and his previous papers cited therein).
The topic of the evolution of birds always interests NCSE readers, and so it was not surprising to find so many good questions in our mailbox that we thought it would be interesting to collect them all together and respond in a single column. First, however, I have to apologize to readers and thank John Ruben of Oregon State for pointing out a misstatement on my part. I said that some dinosaurs such as Beipiaosaurus and Sinosauropteryx had beta-keratins in their integumentary coverings (RNCSE 2000; 20 [5]: 28-31). Sorry! Rather, the alvarezsaurid dinosaur Shuvuuia, which is close to oviraptorids, has beta-keratin in the filamentous, featherlike integumentary structures on its head (Schweitzer 2001). More importantly, like birds and unlike other reptiles, these structures lack the alpha-keratins found in reptile scales and all vertebrate skin. Beipiaosaurus and other feathered and "fringed" non-avian dinosaurs are in the same clade of birds and closely related theropod dinosaurs, but, contrary to what I noted, they have not yet been tested for keratins.
They have the same kinds of integumentary structures, which appear to be homologous to the feathers on today's birds as well as to those of ancient birds such as Archaeopteryx and Confuciusornis. Because all these animals have the same kinds of structures, and they are in the same clade of closely related animals, we infer that all these structures lacked alpha-keratins, and that hypothesis must be tested by future discoveries. I apologize for creating confusion: I was thinking of one animal and writing about another, and my fingers got ahead of my brain (or vice versa). For more information on feather biochemistry and evolution see Brush (2001 and his previous papers cited therein).
Readers Ask
What is a bird in the first place?
Any taxon, or group of organisms, has a definition and a diagnosis. The definition is expressed as a function of descent, or relationship. For example, most of us (but see papers in Gauthier and Gall 2001) define Aves as living birds, plus Archaeopteryx and every animal that evolved from the common ancestor of Archaeopteryx and living birds. This would include extinct forms such as Confuciusornis and Hesperornis (Cretaceous birds that are more distantly related to living groups) as well as birds such as the moa and the dodo (which are within the group of living birds).The diagnosis of a taxon such as birds proceeds from the definition, in part, and it comprises the diagnostic features that were present in the common ancestor of the group as it is defined. What features did that common ancestor have that its relatives outside birds did not? These are the diagnostic features. To reason this out, we do not have to find the exact common ancestor. Rather, we infer from these features what the most basal ("primitive") members of the bird lineage shared that non-birds do not share.
As you can imagine, it is more difficult to give birds the easy diagnosis of "feathers," now that we have full-blown feathers on dinosaurs that are not birds but are closely related to them (and these feathers, like bird feathers, lack alpha-keratins). However, Archaeopteryx and the other basal birds had a full complement of flight feathers, with primary and secondary feathers, and they also had arms that were approximately as long as their legs, unlike their non-flying feathered dinosaurian relatives. There are other more technical features of the skull, vertebrae, girdles, and limbs that separate birds from other dinosaurs (see Padian and Chiappe 1998).
How did feathers evolve?
We are getting closer to a fuller understanding of how feathers evolved. The simplest structures that seem to be related structurally to feathers are seen in the dingy, furlike, thick covering of Sinosauropteryx and its relatives. The first specimens discovered were not preserved well enough to reveal what later discovered specimens did: that these structures were branched (Xu and others 1999). Moreover, there are several kinds of feathers, in addition to these fringe-like structures, on some related non-avian dinosaurs such as Caudipteryx and Protarchaeopteryx (Xu and others 2001; Padian and others 2001). Some of these are flattened, with vanes composed of highly aligned barbs, like typical bird feathers of today. Others have a more down-like aspect, and still others seem to be almost brush-like, gathered at the base into a distinct stalk. Archaeopteryx, the first known bird, preserves no evidence of the latter two types, but has fully developed flight feathers. Many beautifully preserved specimens of the early Cretaceous bird Confuciusornis seem to have a "halo" of feathers that are not flight feathers, but rather look like simpler filamentous structures. And the development of actual feathers has much in common with this evolutionary transition from branched, filamentous structures to vaned, barbed feathers. Of course, we have no records of the actual developmental patterns involved in these ancient structures, and there are many differences between them and the feathers of living birds, too.Don't other extinct animals have feathers?
The bizarre integumentary structures of the (presumed diapsid) Triassic reptile Longisquama have recently been likened to feathers (Jones and others 2000), and have even been called "para-feathers" (Maderson and others 2001). They are long, frond-like structures, but their resemblance to feathers is superficial at best (Reisz and Sues 2000, 2001). They appear to have a central stalk with intermittent, internal pith-like structures superficially like feathers (and also to some plant stems). However, the vane-like structures on either side of the stalk appear to be solid, with wavy corrugations that lack barbs, and the whole thing is surrounded by a discrete band of tissue. The entire structure is not planar but three-dimensional. Nothing of their composition is known, and only one specimen is adequate to provide good details. For these reasons, although Longisquama has been known for over 30 years, no one has considered its structures feathers or feather-like until recently, and this claim has not met much acceptance – to the contrary, there is considerable skepticism. Most qualified observers agree that the similarities to feathers are merely superficial. Because, as I noted in my previous piece, there is so little support for this hypothesis, Longisquama cannot be claimed to have feathers. Moreover, if these were feathers, birds should be closely related to Longisquama, but no one claims this or offers any additional evidence in support. So, if there is no relationship to birds, how can these be feathers?How do we know that the feathered Caudipteryx and Protarchaeopteryx are non-avian dinosaurs instead of secondarily flightless birds?
Good question, especially since they share some morphometric features with true birds (Jones and others 2000). They could indeed be flightless birds, but phylogenetic analyses of the entire range of skeletal features (as opposed to a few morphometric similarities) do not imply this. Caudipteryx is considered related to the oviraptorids by several independent studies (reviewed in Sereno 2001). Isolated morphometric features are interesting, but they have to be assessed as part of a phylogenetic analysis before we can use them to help to determine relationships. Sometimes these similarities are misleading. For example, hummingbirds have unusually short wings, even for their size. Their backs have not become longer, though, and we know this because we can compare a lot of different measurements in the skeletons of other birds related to hummingbirds to demonstrate that it is the wings that have shortened. We also have an explanation: hummingbirds produce lift on both upstroke and downstroke, which are far more symmetrical actions than in other birds. Like any features, morphometric ratios can change, often for very different reasons. We must examine the context of the morphometric difference in order to use it as a character in phylogenetic analyses.What if these specimens are all faked?
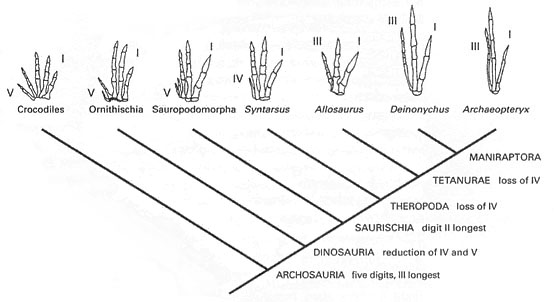
Indeed. Was not National Geographic embarrassed by the specimen of "Archaeoraptor" that turned out to be a mixture of two or more animals assembled by the Chinese villagers who sold the specimen? Yes, but the specimen was still being studied by computerized tomography (CT-scan) to check just this question when National Geographic went ahead with its report. The specimen was never described scientifically, the artifice was quickly discovered (Rowe 2001), and part of the find became the new small dromaeosaur Microraptor (Xu and others 2000). For details on the history of this specimen, see Padian (2000). Tampering has come to be almost automatically suspected in many cases of specimens discovered from certain areas, because they are collected and prepared by local people who know that more complete or more attractive specimens will bring more money when sold to commercial fossil dealers. When there is doubt, the pieces are disassembled, more closely examined microscopically, and CT-scanned if possible. Usually this happens only when there is reason to suspect that some features are unexpected or out of proportion.Birds are said to have fingers that represent II, III, and IV of the pentadactyl hand, whereas theropod dinosaurs have I, II and III, so they cannot possibly be related, can they?
Some inferences from limb development have been used to support this conclusion (Burke and Feduccia 1997). However, no bird embryo appears with the developmental precursors of all 5 fingers (and not simply the cartilage condensations associated with the developing digits). Unfortunately the remaining 4 fingers do not come with numbers on them, so this evidence is inconclusive. Sadly, we have no prospects of developmental series from Mesozoic dinosaurs. What we do have, however, is a robust phylogeny of archosaurian reptiles (Padian and Chiappe 1997, 1998; see accompanying figure) that shows (on the basis of trees constructed from many other characters) how the hand was reduced from a 5-fingered ancestor. In all archosaurs (including crocodiles) the 4th and 5th digits are smaller than the other two. In the first dinosaurs this is still true; in theropods the 5th is at first reduced to a nubbin (Dilophosaurus) and then lost, and the 4th is only represented by the metacarpal (palm bone). This too is lost in all more derived theropods, in which only I, II, and III are preserved. III is lost in tyrannosaurs, and both III and II are lost or greatly reduced in alvarezsaurids such as Mononykus. What is interesting is that the three digits of Archaeopteryx and the other birds preserve the exact forms, proportions, numbers of phalanges, and wrist connections as those of the three-fingered theropod dinosaurs. Coincidence? Not likely.However, there may be a resolution even to the embryological problem. Wagner and Gauthier (1999) suggested that a mutation may have shifted the embryological II-III-IV series in theropods to a I-II-III position such as we see in living birds. A plausible mechanism, based on other developmental examples; it remains to be seen whether tangible evidence in this case will emerge.
What's the take-home message?
Glad you asked. We often encounter creationist arguments that science cannot be authoritative because scientists disagree about the answers to many major questions. Science does not command unanimity or even consensus; as scientists, we do not have to (and should not) accept conclusions that we do not understand or have no expertise in. Though science does not demand consensus, the fact is that science does produce consensus – not by commanding it, and not by ignoring important discrepancies among positions, but by working hard to answer unanswered questions and find conclusive evidence. Most of all, science works by conducting scientific business out in the open in the forum of peer-reviewed scientific research literature. The consensus is not forced but persuaded. However, there is no rule that scientists have to agree. A corollary is that the fact that not everyone agrees on a conclusion does not mean that the conclusion is incorrect. We like to say that science is open-minded but not empty-headed. Contrary ideas and opinions can be held; no vote is taken, no censure given to those who disagree.However, as conclusions become stronger, the bar becomes higher for those who wish objections to be taken seriously. They have more evidence to explain away. They have to come up with a better hypothesis that explains all that evidence and more. And they have to show us that they are using methods of science that the community of scientists in that field accepts. I made the argument that in the case of the origin of birds, the vocal opponents (who actually are far fewer than their representation in the "even-handed" media suggests) do not treat the vast majority of evidence that birds evolved from theropod dinosaurs; they have no rival hypothesis to be tested; and although they do not use cladistic analysis (which is standard for all branches of systematic biology), they do not provide or justify a better method. This debate, which began scientifically in the modern sense in 1973, has gotten to the point where it is no longer scientific, for the reasons just noted.
Many creationists use a literal reading of the Bible as a litmus test for scientific pronouncements. This reading is their authority in all matters, and so it is sometimes difficult for them to understand how science can be authoritative if it is not authoritarian (that is, if there is no governing body deciding the validity of scientific conclusions). How can scientists disagree about something this important and expect us to respect their work? As I noted previously, the scientists involved in this debate do not disagree that birds evolved from reptiles; the questions are which reptiles and how. However, not all views are equally supported by evidence or equally justified by methods. In 1971 we could easily say that the origin of birds was an open question; this is much more difficult to say in 2001. On the other hand, most scientists from many disciplines used to think, on the basis of good evidence, that there was no way that the continents could move.
Thanks again to John Ruben, Brent Warner, Steve Hunter, Alan Brush, John Hutchinson, and other correspondents for good questions and information.