Introduction
Creationists oppose the idea that species can evolve indefinitely and charge evolutionary biologists with failing to define their terms properly. In this article I want to trace briefly the history of the idea of species and show that it is in fact a virtue of biology that it tries to make its terms follow the evidence rather than to define them all up front. The idea that species were universally thought to be fixed prior to Darwin is simply wrong — many creationist thinkers of the classical period through to the 19th century thought that species could change. The issue of evolution was, in fact, impossible to suggest until the claim was made that species were fixed, and as soon as it was suggested, so too was evolution. There has been a longstanding vagueness about living "kinds" that goes back to the classical era and that follows from good observation. What is more, nothing in the biblical or theological traditions requires that species are fixed, only that kinds exist, which neither evolutionists nor traditional creationists ever denied.
Because the number of species "concepts" in the literature is high, I have also tried to put them into context and list them for easy reference (see this page). That way, when a "concept" is referred to in a text, it can be compared to other candidate conceptions. It is clear to me, at any rate, that there are many conceptions of species, and that biologists use the one that best suits the organisms they study. I think of this as a "conceptual delicatessen" — when scientists need a species concept to suit the organisms being studied, they will typically assemble a custom "club sandwich" from previous ideas. This is not bad practice — if science is about learning and using words to express that learning, then we should expect that they would do this, and in fact they should.
To understand the concept of species, we must understand how the notion developed in the history of biological research. One point that must always be borne in mind: people did not suddenly become smart upon the publication of On the Origin of Species, nor were they bad observers before that date. And keep in mind a related point: religious commitment had little to do with the sorts of conclusions natural historians and biologists reached before Darwin, and it seems that in science, it still does not now.
The evolution of the species concept
Then God said, Let the land produce vegetation: seed-bearing plants and trees on the land that bear fruit with seed in it, according to their various kinds. And it was so. The land produced vegetation: plants bearing seed according to their kinds and trees bearing fruit with seed in it according to their kinds. And God saw that it was good. …
And God said, Let the water teem with living creatures, and let birds fly above the earth across the expanse of the sky. So God created the great creatures of the sea and every living and moving thing with which the water teems, according to their kinds, and every winged bird according to its kind. And God saw that it was good. …
And God said, Let the land produce living creatures according to their kinds: livestock, creatures that move along the ground, and wild animals, each according to its kind. And it was so. God made the wild animals according to their kinds, the livestock according to their kinds, and all the creatures that move along the ground according to their kinds. And God saw that it was good.
Genesis 1, verses 11–2, 20–1, 24–5, New International Version
So long as people have been farming plants and raising livestock, they have been aware that one organism gives birth to another very like it. That is, they have known that living things come in kinds. This is not confined to the Bible, of course. Aristotle knew it. So did Theophrastus, his student, sometimes called the father of botany. It is not, as they say, rocket surgery.
So given the relatively short time scale of human observation, it followed that people would tend to think that species, living kinds, were stable; and they did. But they did not think species were unable to change for a very long time, not until John Ray, a brilliant English 17th-century botanist who compiled the first complete flora (of Cambridgeshire, and then of England), wrote in an influential work:
After long and considerable investigation, no surer criterion for determining species has occurred to me than the distinguishing features that perpetuate themselves in propagation from seed. Thus, no matter what variations occur in the individuals or the species, if they spring from the seed of one and the same plant, they are accidental variations and not such as to distinguish a species … Animals likewise that differ specifically preserve their distinct species permanently; one species never springs from the seed of another nor vice versa.
This was the first recorded biological definition of "species", although the logical term had been used in biological contexts for a long time prior to that. But his was not the traditional view. Following a suggestion of Aristotle that new species were formed by hybridization at water holes in Africa, St Augustine, among others (including one of the translators of the King James Bible), happily accepted that new species could be formed out of old ones. Linnaeus himself, who is sometimes regarded as the originator of species fixism, observed hybridization between two plant species in his own garden, and late in life revised his view that species were as the "Infinite Being" had first created them. Certainly there was no tradition in Christian theological circles that species had to be unchanging before then.
During the Middle Ages, little natural history — or biology as we would now call it — was being done. But there was an exception: the Holy Roman Emperor Frederick II of Hohenstaufen (1194–1250) was a keen falconer, and wrote, literally, the book on it, finding that Aristotle was sometimes a bit too credulous, and worse that he failed to discuss hawks, falcons, and hunting birds. Frederick had the resources and the time to do a proper study, and he found that bird species were not simple things at all. He settled on interbreeding as a standard. Albert the Great, who had access both to Frederick's falconers and writings, followed this idea. But both still took seriously enough the old idea of spontaneous generation of species from other species to investigate it. Frederick sent envoys to Sweden looking for evidence for or against the idea that the barnacle goose arose out of worms (which is how it got its name). He found no evidence and concluded that the idea was based on ignorance. Albert did breeding experiments and managed to show that the geese laid eggs in the usual manner.
So we should first of all abandon the idea that people before Darwin thought that species were fixed, necessarily. Some did; many did not. Moreover, almost as soon as the idea of species-fixism caught on, it was challenged. Linnaeus made the idea popular in his Systema Naturae, the first edition of which was in 1735. In 1745, physicist Pierre Maupertuis argued in his Physical Venus that species did evolve, that they did so through a crude version of natural selection, and that inherited characteristics were passed on in a 3:1 ratio through both mother and father. While this did not influence many people at the time, it indicates that fixism was not universal among scientists even when it was still a new idea.
Why is it that we tend to think pre-Darwinians were all fixists? In part this is because Darwin has been used as a turning point in modern biology, which of course he was, and so some, the leading evolutionist Ernst Mayr among them, have tried to make him the ultimate source of all that is correct in modern biology. Furthermore, we think that religious belief before Darwin must have forced people to be fixists. But many quite orthodox Christians held to transmutation of species, and in some cases where this was denied, such as by the great anatomist Baron Cuvier, it was not for religious reasons but from a lack of evidence (although Cuvier managed to present the evidence that florae and fauna were not constant through time, even in Europe).
It is also not true that belief in creation as such forced a species-fixist position. Apart from deism, which perceived God as a creator who effectively left the world to run by the laws he created, many Christians held that the work of creation was still under way. And Christians who were natural historians, whether botanists or zoologists, often described species fairly well.
There is another myth — that before Darwin naturalists thought that species were defined by their morphology or their "essence". But morphology was used by taxonomists simply as a way to identify species, not as the cause of them, and even Linnaeus knew that his "Natural System", as it came to be called, was a useful convention, not a natural system at all. Taxonomists argued about how to define species, but in nearly all cases this was about how many and what kinds of characters were reliable. In the early 19th century, there was no "species problem", but only a "species question", which a minor geologist by the name of Charles Lyell called "that mystery of mysteries" — why were there species? It was not a question he, an orthodox Christian, thought could be answered from Scripture. Neither did his eventual disciple Darwin think that.
Darwin's view of species has likewise been misunderstood, in part because he did not really consider the definition of a species to be the primary question. Like many professional taxonomists (Darwin wrote the first and still one of the best descriptions of barnacles), he found the constant squabbling about whether this variety or that was a separate species or the same to be a nuisance to doing the work. He cited with wry amusement one taxonomist, Phillips, who declared "at last I have found out the only true definition — ‘any form which has ever had a specific name'!" And there was, in 1842, a set of standards from which all modern taxonomic rules derive — Darwin was a committee member — which formally instituted the rule of Linnaeus that species had to have a binomial (a genus and species name), and that only professionals could name species (to stop bird-watching enthusiasts naming every different plumage as a species). In the Origin, he wrote:
… it will be seen that I look at the term species as one arbitrarily given, for the sake of convenience, to a set of individuals closely resembling each other, and that it does not essentially differ from the term variety, which is given to less distinct and more fluctuating forms. The term variety, again, in comparison with mere individual differences, is also applied arbitrarily, for convenience's sake.
On the basis of this and other comments, he seemed to be saying that a species was not a real thing, but that it was just what we called something for convenience. But in his works overall, he treats species as real things, mostly (but not always) isolated by infertility, with different ecological adaptations. His point was, and it remains a sticking point today, that the difference between a species and a variety within a species was vague. This, of course, is due to the fact that species, like sand dunes, rivers and clouds, have no hard and sharp boundaries between them because of evolution.
About the time evolution had been universally accepted by naturalists (now called biologists), but before the new Darwinism of the synthesis of genetics and evolution had been settled, one EB Poulton wrote a paper in 1903 entitled "What is a species?" in which he addressed what now became the species "problem". This set the agenda for the next century. From being the useful identification of kinds that might vary, in the late Middle Ages and after, through to being a problem of who got to name species and how they were to be differentiated, now species were the "units" of evolution, and of biology in general. And a veritable explosion of attempts to define species followed. By the end of the 20th century, there had been some 22 distinct concepts identified by RL Mayden, and depending on how one divides them, some few others have been added. By my count, there are around 26 concepts (see sidebar, p 42–3).
Well, not exactly concepts. There is only one concept, which we label by the word "species". There are 26 or so conceptions, or definitions, which we define in other ways. This slightly picky philosophical point matters. We are arguing over the best way to define a concept. This depends on scientific data, theory, and other factors (some of them political, within the scientific community). We might deny that the concept even has a useful definition, or we might think that we have been misled by the use of a single word and seek a number of different concepts that serve the purposes of science and knowledge. I mention this because one of the oft-repeated claims made by anti-evolutionists is that if we cannot define our terms, we cannot show that species evolve. This canard goes back to Louis Agassiz, the famous geologist and paleontologist, who single-handedly introduced America to biology. Agassiz wrote:
[I]f species do not exist at all, as the supporters of the transmutation theory maintain, how can they vary? And if individuals alone exist, how can differences which may be observed among them prove the variability of species?
Darwin rightly snorted to Agassiz's one-time student Asa Gray:
I am surprised that Agassiz did not succeed in writing something better. How absurd that logical quibble — "if species do not exist how can they vary?" As if anyone doubted their temporary existence.
Creationists will often claim that they are not interested in the species level, though. Initially, creationism did require fixity of species. In the 1920s, when George McCready Price equated "species" to the biblical "kinds", he was forced, to allow for the Ark to carry "every kind", to raise the bar higher. Even this was not original. In the late 18th century, Buffon, Cuvier's predecessor, had suggested that there was a "first stock" from which all members of a kind had evolved, so that all cats evolved from an original animal, modified by geography and climate, for instance. So creationists themselves have a "vagueness problem" no less than evolutionary biology does. Life is vague. Certainly the creationist "kind", or "baramin", as they mangle the Hebrew for "created kind", is extremely elastic. Given that elasticity, the motivation for the inference that was made naturally during the 17th and 18th centuries that species do not evolve is undercut. If kinds are not exact in reproduction, why think that the Genesis account is enough to prohibit evolution? The answer is, of course, that biblical literalism is not the primary motivation here for opposition to evolution.
The species problem
Reproductive isolation conceptions
It begins in 1935, when a young fruit fly geneticist named Theodosius Dobzhansky published a paper "A critique of the species concept in biology" in a philosophy journal. Not that there had not been developments after Darwin. Various people had suggested that species were "pure gene lines" or "wild-types" that did not vary much. Mendelian genetics caused a lot of debate about species. Dobzhansky claimed that a species was:
… a group of individuals fully fertile inter se, but barred from interbreeding with other similar groups by its physiological properties (producing either incompatibility of parents, or sterility of the hybrid, or both).
This was the original genetic version of reproductive isolation concepts (Buffon had proposed interbreeding as a test a century and a half earlier, which Darwin rejected). Unfortunately, a version framed by Ernst Mayr got called the "biological" species concept, in contrast to what were seen as "nonbiological" concepts that relied largely on form and based in museum taxonomy, which were called "morphological" concepts by Mayr. But I think it is better to call these Reproductive Isolation Species Concepts (RISC) than "biological" ones, for any decent species conception is biological. Mayr's version changed over the years, but the one taught to most undergraduate biology students is the original:
A species consists of a group of populations which replace each other geographically or ecologically and of which the neighboring ones intergrade or interbreed wherever they are in contact or which are potentially capable of doing so (with one or more of the populations) in those cases where contact is prevented by geographical or ecological barriers.
Or shorter:
Species are groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups.
Much of the focus on species after this centered on Reproductive Isolating Mechanisms, or RIMs for short. Mayr's view was that species are formed when part of the species is geographically isolated from the main range and evolves in its own way such that when it gets back in contact, RIMs have evolved, as it were, by accident, and the two no longer interbreed successfully. Selection against hybrids, which are, so to speak, neither fish nor fowl in ecological adaptations, then strengthens the isolation (a process called "re-inforcing selection"). Mayr's version of the origin of species, published in 1942 and reiterated for the next 60 years (Mayr survived to 100, outliving many of his adversaries, and thus getting the last word), is called the allopatric theory of speciation. Allopatry means that two populations, or species, or groups, of organisms live in different areas (allo- = other, patria = homeland). The alternative kind of speciation, which is in effect Darwin's view, is called sympatric (sym- = together) speciation, and it is highly contentious among specialists, with some thinking that it occurs, particularly among fruitflies and lake-bound fishes, where it has been studied, and others thinking that it does not, and the debate goes on. It requires that RIMs evolve in place, so to speak, and the naysayers think this is unlikely to occur. If sympatric speciation does occur, then there can only be one reason — natural selection. Recent theoretical work shows that it is possible if the conditions are right. What we do not yet know for sure is how often the conditions are right.
There is another uncontested class of speciation processes — usually involving hybridization, that old idea of Aristotle. In plants particularly, but also in animals, fungi, bacteria, and so on, sometimes entire genetic complements can double, triple or more, resulting in a condition known as polyploidy. When this happens, sometimes the chromosomes and genes do not line up due to differing genetic structures of the parents, but an extra doubling of the genome, followed by a cell division, can give the cell a paired set of chromosomes, allowing it effectively to found a new species in one or a few generations. When two species interbreed, this allows the resulting organism to have a matched set of chromosomes. It has been estimated that nearly all ferns, for example, have a case of polyploidy in their ancestry, and as many as 7% of actual fern species are formed this way. It has also been seen in flowering plants, corals, grasshoppers, other insects, and reptiles. It is even hypothesized that the entire mammalian branch of the evolutionary tree was started with this kind of event. So in a sense, Linnaeus and Aristotle were right … sort of. Even Mendel thought this might be the reason why new species evolve, which informed his research into hybrid forms, although he studied hybridization within, not between, species.
Evolutionary conceptions
So the RISC conceptions have a lot of leeway for interpretation. But they are not the only conceptions on the board. One kind of conception goes by the name evolutionary species concepts, which is also a misnomer (because all species have evolved). On this view, it does not much matter if two possible species under consideration are reproductively isolated. Even if there is gene flow between them on a regular basis, what counts is whether or not they remain evolutionarily distinct. Dobzhansky's definition has a hint of this, but the original formulation is due to a paleontologist, George Gaylord Simpson. Simpson defined it this way in 1961, although there is an earlier and more technical definition from ten years before:
An evolutionary species is a lineage (an ancestral–descendant sequence of populations) evolving separately from others and with its own unitary evolutionary role and tendencies.
What counts here is that no matter what happens in terms of gene exchange, the populations remain distinct, and have their own forms, adaptations, and fate. The term "lineage" used here is particularly important, as it focused biologists' thinking more in evolutionary terms, and gave rise to yet another class of conceptions — phylogenetic species concepts.
Evolutionary conceptions have been expanded since Simpson to include asexual organisms (which do not, strictly speaking, form populations, since that term involves interbreeding). The important point is that there is a single lineage over time. In contrast, the RISC conceptions involve a single time "horizon", which means that a species is something that at a particular time and place is not interbreeding with other populations, and they also, necessarily, exclude asexual organisms.
Phylogenetic conceptions
There is a group of species conceptions that go under the shared name of phylogenetic species concepts. A phylogeny is, of course, an evolutionary history, and the initial proposal for a phylogenetic conception came from Willi Hennig, an East German entomologist who nevertheless managed to influence a great many biologists during the height of the Cold War. Hennig's methodology and philosophy of classification is known today as "cladistics".
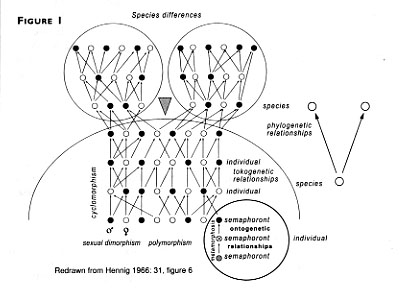
Hennig did not set out to come up with a new species concept. He pretty much assumed something like an amalgam of Mayr's and Simpson's definitions. The difference was that he focused on the lineage element, and combined it with a clear and formal account of making groups logically. In his book Phylogenetic Systematics, translated into English in 1966, he included a diagram about speciation (redrawn above as Figure 1), and when to name a new species.
There are several lineages in this diagram. Ignoring the technical terminology (Hennig was a great one for coining classical names), you can see that each individual organism is part of a genealogical lineage. These, when grouped together, form a species lineage. Clearly what makes a species lineage is the fact that the overall tangled net of genealogical lineages has not yet divided (as in the right-hand diagram marked "phylogenetic relationships"). When it has, says Hennig, the old species is extinguished and two new ones come into being (as in the separate circles at the top).
This provision caused a lot of anxiety. It seemed to be saying that a species has to go extinct when new ones come into being, but of course a new species can evolve without modifying the old one much, if at all. Hennig's convention, as it is known, was more a point about naming species than a definition of their biological nature. When a new species arises, the old name refers to only a part of its descendants and for Hennig that meant it was no longer a "natural" group being named. It is rather like calling rock music "blues" because it is a descendant of blues. What Hennig said would mean that what is still "blues" has to get a new name so that musicologists can talk without ambiguity (so it might get called "traditional blues").
Phylogenetic species conceptions come in three broad flavors. One is the "pure" Hennigian conception described above. But Hennig's methodology also relied on dividing organisms by shared features, which is a diagnostic question. So another phylogenetic conception, which I call the autapomorphic conception (after one of Hennig's technical terms that means "derived forms"), defines a species as the final node on a phylogenetic tree, which is based on comparison of many features. If the analysis does not produce any smaller group, then the organisms in this group constitute a species. This has a side effect of increasing the number of species over the older RISC conception — by as many as five times — and for this reason it is not accepted by many taxonomists. On the other hand, if a species really is just the as-yet-undivided tangled net of genealogical relations, this is what we have to deal with. The autapomorphic conception tends to leave historical issues about whether or not the phylogenetic tree is a good historical representation to one side. The "species" here is diagnostic.
The other conception I call the phylogenetic taxon conception. On this view, a species is just another kind of taxon in a phylogenetic tree — one that happens to be monophyletic and undivided. Monophyly in this case simply means that no descendent of the original species is excluded, so that if a single species did divide into two, it would have been a species before, but is now two species, which is pretty obvious. However, this view allows the old name to be "kept" by one of the descendants.
Phylogenetic conceptions are in a way parasitic on biological conceptions like RISC. In order to know that two organisms are in the same species, one has to eliminate subspecific features such as, for example, different plumages or immune system molecules or genes. Otherwise one can divide the organisms up right down to individuals with novel mutations or slight variations. There are those who do this, too, who intend to eliminate "species" from the working vocabulary of biologists and replace it with "evolutionary group" or "least inclusive taxon"; we may refer to them collectively as "species eliminativists".
Ecological conceptions
In the 1920s, a Swedish botanist named Göte Turesson proposed that there were different kinds of "species", one of which was a response to the local ecology. Though not Mendelian, as Turesson appeared to think the environment changed genes directly, his ecospecies concept has been revived from time to time. Various authors, including Mayr, have suggested that what makes a species is occupation of an ecological "niche", and in 1976 American botanist Leigh Van Valen suggested that a species was a lineage "which occupied an adaptive zone" differently to other species. Van Valen's "proposal" (he did not call it a definition) combined both Mayr's RICS account and Simpson's evolutionary account, for the "adaptive zone" idea is Simpson's as well.
Van Valen's example case was the American white oak complex, Quercus, which will interbreed fairly freely, and yet remains stable phenotypically. This is due, he said, to adaptation for particular ecological needs. Similar cases have been found elsewhere; for example, Australian eucalypts interbreed with more distant relatives, but are often infertile with closer ones.
The idea that underpins this conception is the famous "adaptive landscape". A species, no matter what the gene flow between populations of other species, occupies a "peak" in that landscape (see, for example, the view espoused by Richard Dawkins in his Climbing Mount Improbable [New York: WW Norton, 1996]) where the metaphor of an adaptive landscape implicitly assumes this. Ecospecies are formed when the adaptive landscape "fractures", to use a term from philosopher Kim Sterelny. Recent work on speciation suggests that this is a factor in most cases, although it has a different role in allopatric speciation than in sympatric or polyploid speciation.
A grab bag of conceptions
As with any taxonomy, there are a few things that do not fit neatly into this scheme. Many of the current conceptions mix and match aspects of these conceptions, so a particular description might be characterized as, say, an isolationist, evolutionary, genetic account. But there are also species conceptions that make positive proposals, for example, for asexual species. At one time it was open to doubt that such things existed, or were rare, but apart from bacterial species (which sometimes do not have "sexes" but can exchange genes in various conditions), there are an increasingly known number of "parthenogenetic" species (in animals — in plants they are called "apomictics") that do not need to fertilize their ova or seed. Organisms that are descended from sexual organisms are sometimes asexual, such as the famous whiptail lizards of the southern USA and northern Mexico (genus Cnemidophora). In many cases these are formed by hybridization between closely related species, in animals and plants. Viruses also form "species" called "quasispecies", usually without crossing over their genetic material.
Another species conception is sometimes called "conventionalism", or less accurately but more commonly "species nominalism". This view is very popular among those whose ideas about evolution derive from the work of geneticist JBS Haldane early in the 20th century. It is basically the claim that species are just names, chosen for convenience, so that specialists can talk to each other. However, if species are just convenient fictions, how do specialists know that their terms refer to the same things?
And finally let us consider the problems of identifying species in fossils. Not all information about an organism is recorded in a fossil. We have data about hard parts, and, more rarely, skin or feather impressions, but we are not given their genetics, behaviors, colors, ranges, or mating preferences. And these are often the markers of being a RISC species, or are used to diagnose phylogenetic species. "Paleospecies", as they are sometimes called, may be less arbitrary than conventionalist species, but they do not necessarily map onto "biological" species.
Consider the debate in human evolution over whether Homo sapiens left Africa entire, as it were, or whether a prior paleospecies, Homo erectus, interbred with H sapiens afterwards. Recent work by Alan Templeton suggests there have been three major migrations out of Africa, each of which has left a genetic mark on the modern population. So, are H erectus and H sapiens one species, or two? We may be able to work that out through the sort of work Templeton is doing, but we equally may not.
Knowing species
Now the "biological", the evolutionary and the phylogenetic conceptions have an operational problem for biologists — except in very rare circumstances, it is almost impossible to give the criteria of the definitions for identifying and demarking species. There have been cases where two forms have been identified as the same species because they were observed mating, but there just are not the resources or the time to do experiments of assortative mating to tell in every case. In fact, many "biological" and evolutionary species, such as tigers and lions, which have been isolated for millions of years, can interbreed in artificial conditions, and their progeny are fertile, so mating tests might not help anyway. Back in the 18th century, Buffon established that some species could occasionally interbreed (which is why he thought that there was a first stock for groups of animals). So the definitions, or conceptions as I prefer, are not much help. And so far as evolutionary conceptions go, we simply have no direct access to the evidence we need in the case of extinct and even extant species. It might be true that species have distinct fates, but often we do not know.
It is important to separate the issues of what species are from the issues of how species are known or identified, because while evolutionary biology and genetics indicate some of the processes by which species evolve, the tests we use to find out whether two organisms are the same species or not may in fact be quite incidental to the causes of their being species. Many tests are used, mostly genetic and molecular tests these days. A recent proposal called DNA barcoding uses a particular gene on the cellular organelles known as mitochondria quickly to identify species for conservation and other purposes, but nobody thinks that the gene, COX1, causes speciation. In fact in cases where the "speciation genes" have been studied, they are nuclear genes, not mitochondrial genes (and anyway it is unlikely that the same genes cause speciation in all groups). When creationists attack evolutionary biology for not being able to define the term "species", they are confusing the identification and diagnosis of species and the meaning of the term "species". Occasionally, some scientists do the same thing.
Suppose we take a nonbiological example — "mountain". If we cannot give a universal definition of that term, it does not mean there are no mountains, or even that we cannot tell whether we are looking at one. And the geological theory of plate tectonics explains why there are mountains even when the term is not definable. Definitions are for philosophy, but science can do without them if it needs to. Species are the phenomena of biology that the theory explains, not a priori concepts that have to be clarified exactly. It may be that, as some propose, we ought to replace the term with a range of other terms, such as "evolutionary group", "least inclusive taxonomic unit", and so on. Some "species" will be both evolutionary groups and least inclusive taxa. Some may not. The jury is still out as to the worth of these ideas.
Evolutionary biologist Massimo Pigliucci and philosopher Jonathan Kaplan have proposed that the term "species" is in fact what philosophers call a "family resemblance predicate". This is when being an instance of a general kind referred to by a word (the classic philosophical example is "game") has many criteria, and as long as most of them are met by a particular instance, it is a part of the kind. If evolution proceeds as we should expect from modern theory, then this plethora and confusion of species concepts is easily explained — evolution is a gradual process at genetic and behavioral scales (but it can be abrupt at geological and ecological scales), and so we will see species in all kinds of stages of speciation, extinction, interbreeding compatibility, and so on. On a creationist account, of course, no such explanation is forthcoming. If "kinds" are fixed, we should see only the expression of created variety (and of course that variety could not have possibly passed through the Ark, but let us assume that creationism is separate from Flood literalism), and that should be definite and limited. Instead we see species in groups of varying and ill-defined variety. It is a lot of weight for a simple word — "kind" — to bear. Science does not need it — so long as the ways words are used by each speciality will serve to describe what is observed.